Os Vegetais e a Água
TRANSPORTE NO XILEMA
O xilema forma um sistema contínuo que,
partindo do centro da raiz, atravessa o caule e atinge as folhas, estejam estas a
centímetros ou a muitos metros do solo (Fig. 1 A). Há duas teorias para explicar o
mecanismo pelo qual a água ascende num vegetal. São elas: COESÃO - TENSÃO e PRESSÃO
RADICULAR.

Figura 1A. Fluxo da água no sistema
solo-planta-atmosfera. (Redesenhada de Devlin, 1976)
De acordo com a Teoria da Coesão-Tensão,
a força propulsora para a ascenção da água pelo xilema está exatamente na perda de
água pelas folhas ou, simplesmente, transpiração.
A transpiração consiste na perda de água
pela planta, sob a forma de vapor d’água, que se dá principalmente pelas folhas
(Fig. 1 B)

Figura 1B. A difusão do vapor
d’água na folha. (Redesenhada de Taiz & Zeiger, 1991.
Apesar da grande importância da água para
a planta, aproximadamente 98% da quantidade absorvida é perdida durante a transpiração.
A eficiência da planta está em utilizar mínimos 1 a 2% para a sua dinâmica
metabólica.
A taxa de transpiração depende de fatores
relacionados à própria planta e de fatores ambientais, como umidade relativa do ar,
temperatura, disponibilidade de água no solo, vento e luz solar. Existe uma relação
inversa entre a umidade relativa do ar e a velocidade de difusão do vapor d’água,
de modo que quando o ambiente encontra-se úmido reduz-se a taxa de transpiração. Já um
aumento de temperatura tende a elevar a taxa de transpiração. Assim também age um vento
leve ou brisa, ao retirar o vapor d’água da superfície da folha, acelerando a
evaporação. A luz solar atua aumentando a temperatura da folha, como também estimulando
a abertura do estômato (Fig. 2).
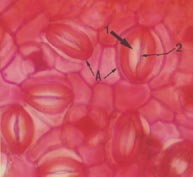
Figura 2 -Vista frontal da epiderme
abaxial da folha de Byrsonima sericea ocorrente no Parque de Pituaçu, Salvador -
BA, evidenciando os estômatos (A) com as células guardas (1) e região de abertura (2).
Dentre os fatores da planta que controlam a
transpiração, o mais importante é o movimento estomático de abertura e fechamento.
A transpiração ocorre preferencialmente
pelos estômatos, presentes em maior quantidade nas folhas (transpiração estomática).
Também através da cutícula, camada lipídica, que recobre as células epidérmicas
(transpiração cuticular) ou em menor proporção pelas lenticelas dos caules
(transpiração lenticelar). Os estômatos são responsáveis por 95 - 97% da água
transpirada, enquanto a participação da cutícula fica, em torno, de 5%.
Ainda que os estômatos predominem nas
folhas, também podem existir em caules jovens, flores e alguns frutos. Quando nas folhas
podem ocorrer em uma das faces (abaxial ou adaxial) ou em ambas.
Os estômatos são formados por duas
células especiais denominadas células guardas ou oclusivas, que apresentam paredes
diferentemente espessadas. Por conta disso, a capacidade de distenção ou de aumentar o
volume das mesmas é bem maior do que a das outras células. E assim com a entrada de
água e conseqüente distenção, essas células se afastam, criando um orifício ou poro,
por onde se difunde o vapor d’água. Com a saída de água, as células-guarda
reduzem o seu tamanho, se aproximam e desaparece o ostíolo e o estômato se fecha (Fig.
2).
Vários fatores como água, luz, CO2
e temperatura influenciam o mecanismo estomático de abertura e fechamento. Normalmente,
quando o solo está bem suprido de água, os estômatos se abrem ao amanhecer, podem
fechar temporariamente nas proximidades de meio-dia e se abrem de novo, à tarde. É certo
porém que para a maioria das plantas, bem antes do por-do-sol, os estômatos já se acham
parcialmente fechados.
O fator que leva à abertura do estômato
é a entrada de água na célula-guarda, ou o seu turgor. Uma das características dessas
células é a capacidade de acumular íons k+ . A tal fato tem-se atribuído,
recentemente, a redução do y de água dessas células, imprescindível ao movimento,
levando a abertura do estômato.
A passagem de K+ para o interior
da célula-guarda se dá por absorção ativa. A energia necessária para esse transporte
vem da hidrólise do amido em acúcar
.As células-guardas também acumulam amido
que é então degradado a um composto, chamado PEP (Fosfoenolpiruvato), pela glicólise. O
PEP então se combina com o CO2 que penetrou pelo estômato, resultando em
ácido oxalo-acético que se reduz a ácido málico. É desse ácido que são liberados os
prótons H+. Com gasto de ATP, esses prótons são expulsos do interior da
célula-guarda em troca de íons K+, que se acumulam. Esse acúmulo de K+
leva à redução do y de água da célula e a um subseqüente movimento de água das
células vizinhas para as células-guarda, que se incham , distendendo-se e ao se
afastarem uma da outra permitem a abertura do estômato.
Há alguns anos atrás pensava-se que a
redução do y das células-guarda era causada pela presença de glicose, produto da
hidrólise do amido e que o CO2 presente no interior do estômato, era fixado
pela fotossíntese realizada pelos cloroplastos das células-guarda. Hoje sabe-se que
esses cloroplastos não convertem CO2 em açúcar.
Numa situação inversa, quando a água
transpirada pela planta supera aquela absorvida, os estômatos se fecham, para proteger o
vegetal contra a desidratação. O fechamento do estômato ocorre porque as
células-guarda perdem água para as vizinhas, numa condição oposta a da abertura. Ao
perderem água, reduzem os seus volumes e em conseqüência disso, desaparece o ostíolo,
e o vapor d’água não passa.
E agora podemos voltar a falar sobre a
condução da água no xilema, pois ao perderem água por transpiração, as células das
folhas reduzem os valores do seu potencial de água. Essa redução de y de água cria uma
pressão negativa ou tensão, que se propaga ao longo da coluna do xilema. Quando as
células foliares retiram água do xilema do caule, esse retira das raízes que retiram do
solo. Forma-se, com isso, uma coluna líquida da raiz a folha, que ascende puxada de cima
para baixo, uma sucção causada pela transpiração.
TEORIA DA PRESSÃO RADICULAR
Opera também no xilema um outro mecanismo,
bem diferente do mencionado acima, e que permite a ascenção da coluna de água a
pequenas alturas. A sua causa está numa pressão desenvolvida pelas raízes, daí a
denominação Pressão de raiz ou Pressão radicular. A magnitude (0,1 Q a a 3 MPa) da
pressão gerada não é suficiente para ascender a água em vegetais de grande porte, por
isso a sua ação se restringe a espécies herbáceas.
A pressão de raiz desenvolve-se quando a
transpiração é baixa ou ausente, o que faz acumular soluto no xilema, reduzindo o seu
potencial de água (y de água) e causando o movimento da água a partir da raiz.
Um fenômeno comum da pressão de raiz é a
GUTAÇÃO, através do qual o excesso de água é eliminado pelos hidatódios (estruturas
similares aos estômatos) sob a forma líquida. Esse fenômeno é mais freqüente à
noite, quando a umidade é alta, o solo está bem suprido de água e a transpiração
está suspensa. Corresponde a gotículas de água que surgem nas margens das folhas de
espécies herbáceas.
E assim concluímos o assunto na certeza da
compreensão do sistema solo - planta - atmosfera, através do qual se estabelece um fluxo
dinâmico da água.
Hermínia Maria Bastos Freitas
E-mail: hbastos@ufba.br
|| Página Inicial | Os Vegetais e a Água | Água no Solo e
Raiz | Transporte de Água na Planta ||